河北省秦皇岛市寒武系张夏组顶部均一石主导的微生物礁
郭翰良1,*, 秦仁月2
(1. 中国地质大学 (北京) 地球科学与资源学院, 北京 100083
2. 贵州乌江能源投资有限公司贵州页岩气勘探开发有限责任公司, 遵义 563400)
摘要: 叠层石、凝块石、均一石以及树形石被并列划分为典型微生物碳酸盐岩, 其中均一石以隐晶质岩性、凝块结构不发育且无明显宏观构造为特征。自从均一石1995年被命名以来, 在地层中很少得到观察与识别, 而且现代类比物也十分缺乏, 因此使得研究均一石具有极其重要的意义。为了解华北地台寒武系的均一石的特征, 选取河北秦皇岛寒武系张夏组构成三级层序的强迫型海退体系域顶部集中发育的一套由均一石主导的、数十米厚的微生物礁, 采取野外采样及室内镜下微观特征观察相结合的方法进行研究。均一石由致密泥晶和少量微亮晶构成, 构成均一石的致密泥晶中可见类似于现代织线菌 (Plectonema) 的丝状体葛万菌 (Girvanella) , 局部可见灌木丛状的附枝菌 (Epiphyton) 以及基座菌 (Hedstroemia) 。研究结果表明, 均一石主导的微生物礁的形成受复杂的有机矿化过程影响或受早期的蓝细菌主导的、发生微生物膜或微生物席内的石化作用影响。因此在了解古老微生物礁的形成过程中, 受蓝细菌主导、经微生物席钙化作用而形成的秦皇岛寒武系张夏组均一石所主导的微生物礁是一个重要的窗口。
关键词: 秦皇岛, 张夏组, 均一石, 微生物碳酸盐岩, 钙化蓝细菌
DOI: 10.48014/cesr.20230702002
引用格式: 郭翰良, 秦仁月. 河北省秦皇岛市寒武系张夏组顶部均一石主导的微生物礁[J]. 中国地球科学评论, 2023, 2(3): 19-31.
文章类型: 研究性论文
收稿日期: 2023-07-02
接收日期: 2023-07-14
出版日期: 2023-09-28
0 引言
希腊语中的单词“leios”的原意是指“均质且平滑的岩石”,后经演变用来表达均一石(Leiolite)这一术语[1,2],Braga J C等[3]在描述西班牙东南部中新世的陆架碳酸盐和硅质混合沉积中的微生物丘时首先将均一石纳入微生物岩的研究范围,并将其定义为“相对贫乏构造而且非晶质的中等构造[2,4,5]”,曾经均一石被认为是“隐秘微生物(Cryptomicrobial)”的同义词并将其归类于凝块石,但后续的研究表明,均一石贫乏组构的特点,使之与早期微生物碳酸盐岩分类中的凝块石和叠层石相异,因此不能将其简单归类。Burne R V等[6]以及Riding R[7-9]等基于中等构造,将均一石作为一个单独分类进行划分并将微生物岩划分为四个类型,其中岩性为隐晶质的、凝块结构不发育且无明显宏观构造的被命名为均一石。
均一石主要见于古老地层中,如Riding R[10]描述的碳酸盐泥丘、寒武系芙蓉统长山组和凤山组地层[2,11,12]以及苗岭统徐庄组[13]中的均一石生物丘。均一石自从1995年被命名以来,在地层中很少得到观察与识别,而且现实中可与之相对比的现代类比物也十分缺乏,在现代的沉积环境中,尽管可以观测到与之有一定相似性的可对比物,如:(1)在巴哈马台地与澳大利亚鲨鱼湾中的由碳酸盐沉积物构成、缺乏纹层与凝块结构的特别的“叠层石”体;(2)咸水湖中的碳酸盐岩结壳;(3)具碎屑结构的微生物沙丘等[2],但与古老地层中的均一石相比仍有一定差异。因此对均一石的沉积组构、成因机制以及形成环境的研究具有极其重要的意义。
寒武纪作为显生宙第一个纪,其独特的沉积作用特征和独特的碳酸盐沉积样式产生了诸多影响,包括:由于微生物的活动增加,产生了大量与之相关的灰泥;由于后生动物辐射,微生物碳酸盐岩的丰度也呈现出正相关的增多;“显生宙早期第一幕蓝细菌钙化作用事件”[14]发生;寒武纪早期,由于生物造礁建设系统的崩盘,使得寒武纪海洋普遍呈现“贫乏骨骼”[15]。
本次研究的河北省秦皇岛市寒武纪苗岭世张夏组构成三级层序的强迫型海退体系域顶部,存在一套由均一石主导的微生物礁,其中发现了明显的微生物代谢活动和钙化的证据,生物礁的致密泥晶中发育着具有多种表现形式的钙化蓝细菌,其中含有葛万菌类(Girvanella)、附枝菌(Epiphyton)以及基座菌(Hedstroemia)等,是了解显生宙早期第一幕蓝细菌钙化作用事件[14]的关键现象,也为进一步了解均一石如何形成于复杂的微生物膜的代谢以及钙化作用、如何诱发的碳酸盐原地沉淀作用提供一个较为典型的实例和重要材料。
1 地质背景
1.1 构造与沉积背景
华北地台属于典型的陆表海浅水沉积盆地[16],是我国重要的构造单元之一。研究区(图1a)地处于我国华北地台东北缘,秦皇岛市北部山区,距市区约30km(图1b),处于郯庐断裂带的西部。研究区出露新太古代花岗岩,长城系、蓟县系和青白口系地层在之上沉积。研究区地层厚度变薄,长城系碳酸盐岩沉积比例较天津蓟县中、新元古界标准剖面明显减少,其中碎屑颗粒物质的粒径较粗、存在铁岭组、洪水庄组和下马岭组部分层位缺失,几大沉积层序表现出明显的古陆边缘特征以及典型的底界超覆现象[17]。
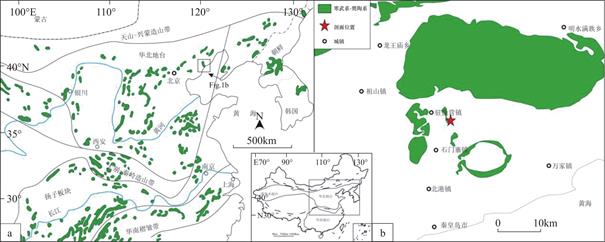
图1 华北地台寒武系的露头分布
Fig.1 Distribution of Cambrian outcrops on the North-China Platform
1.2 层序地层划分
华北克拉通形成于1850Ma左右,此后大部分地区发生隆起,形成古陆,而裂陷谷盆地和被动陆缘盆地则广泛出现在其北缘、西缘和南缘[18],研究区出现沉积厚度巨大、沉积序列良好的浅海相碳酸盐地层。骨骼在寒武纪还尚未成为海洋中的碳酸盐沉积物的重要沉积来源,故该时期风暴海洋的特点是“贫乏骨骼的”[13,20,22]。研究区整体发育的退积序列如图2所示,由老至新依次为第二统、苗岭统以及芙蓉统的沉积物,其中第二统为红层与碳酸盐岩混合沉积物,苗岭统为鲕粒滩碳酸盐台地序列,芙蓉统以灰泥为主的碳酸盐台地沉积序列[19,20,23]。秦皇岛地区的苗岭统由混合潮坪沉积体系的毛庄组和徐庄组与缓坡型台地沉积体系的张夏组和崮山组共同组成,崮山组、张夏组与徐庄组的接触关系为淹没不整合接触,其间张夏组和崮山组上部分布有大片的鲕粒滩,自下往上组成一个沉积变化序列[20]。在张夏组沉积时期,华北地台受海侵作用影响最大,主要包括两个相带,其中西北小部分为夹杂在环潮坪相中的古陆,其余部分内部为环潮坪相,代表着毛庄组和徐庄组下部的以白云岩为主的混合潮坪沉积体系;外部为中至缓坡相(图3),代表着张夏组和崮山组上部的以泥晶灰岩为主的缓坡型碳酸盐沉积序列。
秦皇岛的寒武系苗岭统张夏组构成完整的淹没不整合三级层序[13,14,19,20,22],崮山组与徐庄组的岩性为具有凝缩性质的中至深缓坡相钙质泥岩夹泥灰岩(图2),地层直接覆盖在鲕粒滩相灰岩地层之上[13,19,20]。在张夏组顶部的强迫型海退体系域[24]的顶部发育有一套鲕粒滩相泥晶灰岩,其中的叠层石、均一石生物丘与之共同构成潮下型米级旋回,代表较高的碳酸盐沉积和较慢的侵蚀作用下的高能沉积单元。
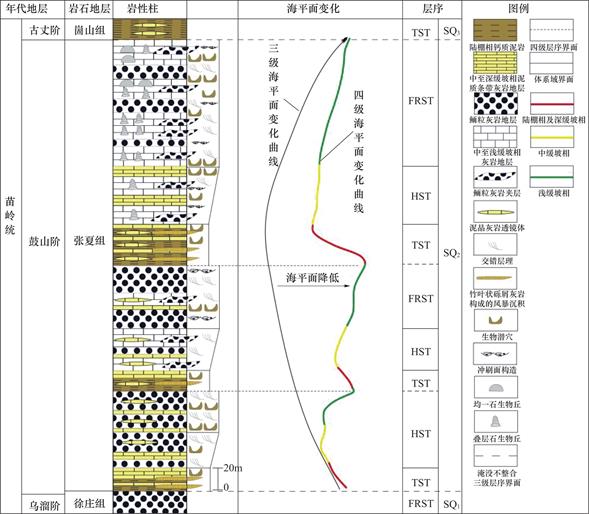
图2 秦皇岛寒武系张夏组地层柱状图(修改自参考文献[21])
Fig.2 Stratigraphic histogram of Zhangxia Formation of Cambrian from Qinhuangdao City(Modified from Ref.[21])
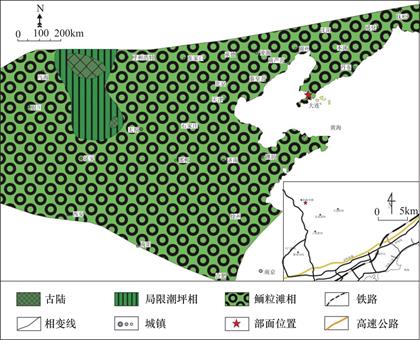
图3 华北地台寒武纪苗岭统张夏组沉积期晚期相带分布(修改自参考文献[19])
Fig.3 Depositional zonation of the last sedimentary period of Zhangxia Formation of Miaolingian in Cambrian,North-China Platform(Modified from Ref.[19])
2 实验方法
2.1 野外采样
本次研究的采样点位于河北省秦皇岛市北部山区,距市区约30km,所采样品为寒武系苗岭统鼓山阶张夏组的泥晶灰岩。
2.2 室内显微镜观察
2.2.1 岩石薄片制作
本次研究过程中使用的岩石薄片制作标准遵循中华人民共和国石油天然气行业标准《岩石制片方法》(DZ/T 5913—2004)。
2.2.2 显微镜型号
本次研究过程中室内镜下观察所使用的显微镜型号为蔡司Axiolab 5,使用透射光和正交偏光相结合的方法进行观察。
3 结果
3.1 均一石生物丘的宏观特征
如图4a所示,河北秦皇岛均一石生物丘集中产出于张夏组顶部,直接覆盖在叠层石生物丘之上,两者有明显的产出差异且有一条较为清晰的界限(图4b),且与崮山组底部也有一条清晰界限(图4f)。由均一石主导的微生物礁表现为灰白色致密均一的块状泥晶灰岩,贫乏纹层和凝块状或树枝状组构,可见冲蚀沟槽(图4c),冲沟内充填着细小颗粒组成的灰岩,界线比较清晰,代表泥晶灰岩组成的均一石和颗粒灰岩共生的特征,可能代表较强的风暴流或水流的改造和破坏作用,表明该微生物礁形成于动荡高能水体环境中固着底栖沉积物的微生物席或微生物膜内的碳酸盐原地沉淀和钙化。由均一石主导的微生物礁局部可见类似于凝块石和树形石中特征性的“凝块”(Clot)和树枝体(Dendroid)组构,表现出“凝块石(Thrombolite)”或者介于树形石和凝块石之间的“树形状凝块石(Dendrolitic thrombolite)”[25]的特点,姑且称之为“凝块状均一石”或“树形状均一石”,但树形石与均一石整体上是呈现过渡变化的,不存在一个非常清晰的界面或者界线,往往是由“均一石”逐渐演变为“树形石质的均一石”“均一石质的树形石”再到典型的“树形石”。均一石的局部可见泥晶灰岩充填的、代表后生动物居住的生物遗迹(图4d),还可见选择性白云石化特征的生物潜穴(图4e,4f),成因可能是造穴动物有机基质中的异养细菌降解作用的产物[14,26],抑或为成岩微环境中受到高含量有机物影响而产生的白云石沉淀物[27]。

图4 张夏组顶部均一石主导的微生物礁宏观特征
Fig.4 Macroscopic feature of the microbial reefs predominated by leiolites at the top of Zhangxia Formation
3.2 均一石生物丘的微观特征
华北地台寒武系发育在古生代第一次海侵背景下,在华北地台大部分地区沉积了浅海相碳酸盐岩,海平面的持续抬升有利于碳酸盐的产生和微生物的生长[28]。寒武纪的微生物碳酸盐岩通常出现在碳酸盐缓坡上,其有利的生态位置能够支持框架型微生物岩丘内的不同类型的生物进行生产活动[29],有机体群落聚集在这些生物丘内,并与它们所粘结的沉积物质一起构建了这样极其多样化的生态系统[10]。
秦皇岛张夏组顶部的由均一石主导的微生物礁主要由暗色致密泥晶以及少量亮晶、微亮晶方解石构成,局部可见毫米级别的暗色泥晶凝块(图5a),这些微凝块(图5b是图5a虚线框区域的放大)之间还可见到明显的皮壳光滑的底栖鲕粒[22,30],大小介于0.1~0.3mm,该底栖鲕粒的存在可能说明在富营养状态下,较厚的微生物席内[31]表现为放射纤维状方解石内部的微生物沉淀作用[32]的结果。这些富集有机质的泥晶凝块形态极不规则,在高倍镜下可见内部由葛万菌类的丝状钙化蓝细菌鞘化石构成;均一石致密泥晶组构中还可见三叶虫骨架碎片化石(图5c)和代表氧气气泡残余物的窗格状组构[33,34],这也是光合作用发生的重要标志。均一石还可见较为密集的生物潜穴,潜穴内充填物近乎完全白云石化,形成菱形的雾心亮边白云石晶体,可能代表着受某些有机质含量高的成岩微环境影响而形成的白云石沉淀物[27]和选择性白云石化的产物[14,26](图5d,5e,5f)。均一石的这些微观特征与贫乏构造的宏观特征形成强烈的对比,反映了组成的均一性和不规则的增生作用[11],也说明了并不像曾经认为的那样均一石是完全由致密泥晶构成的。最为特别的是,均一石致密泥晶组构中发育了表现形式多样的丝状钙化蓝细菌鞘化石,其中包括:基座菌(Hedstroemia;图6)、附枝菌(Epiphyton;图7)以及葛万菌(Girvanella;图8),具体描述如下。
3.2.1 基座菌(Hedstroemia)
在构成树形石质均一石生物丘的致密泥晶和微亮晶基质中可见到单株呈房室状的基座菌(图6a,图6b是图6a中虚线框区域的放大),这些菌落可构成泥晶团块(图6c,图6d是图6c中虚线框区域的放大)和毫米级别的基座菌菌落,也可发育在窗格状组构的周围(图6e,图6f是图6e中虚线区域的放大)。菌丝体由泥晶质管构成,直径较细,多在10~30μm之间,呈放射状直立生长,管还可形成分叉树状体,在纵向上呈现扇形态的束状体,其高度一般不超过0.5mm,相邻的管与管之间紧密结合,可能共用同一个的壁或隔膜。现代胶须菌作为固氮蓝细菌中的一个较为典型的实例[8,14,35,36],其管的异型细胞的终端指向菌落中心这一定向排列的特征和呈毛发状的丝状体的放射状排列的特征均和上述基座菌有一定的可类比性。
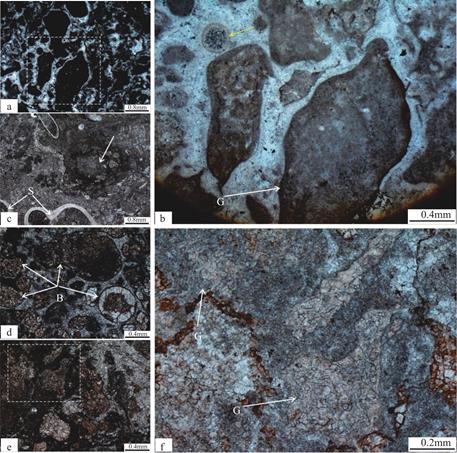
图5 张夏组均一石微观特征
Fig.5 Microscopic feature of leiolites in Zhangxia Formation
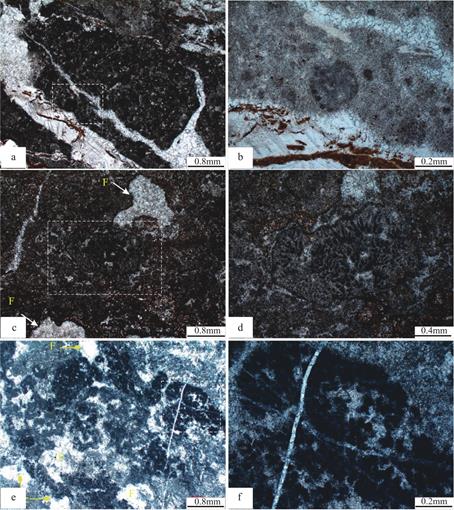
图6 均一石生物丘中的基座菌
Fig.6 Hedstroemia in the leiolite bioherms

图7 均一石生物丘中的附枝菌
Fig.7 Epiphyton in leiolite bioherms
3.2.2 附枝菌(Epiphyton)
关于附枝菌的分类,通常依据其分杈直径及所表现出的菌落基本形态,将其分为4种类型[37,38]:即较粗的枝杈状(类型Ⅰ)、分段枝杈状(类型Ⅱ)、较细的枝杈状(类型Ⅲ)以及管状枝杈状(类型Ⅳ)[39]。秦皇岛张夏组均一石中,不仅有灌木丛状基座菌构成的树形状暗色凝块,还可见成簇生长或单独产出的附枝菌(图7a,7b),菌株呈直立生长的扇形,聚集在一起可形成灌木丛状群落,高度一般不超过0.5mm,并且在一定位置出现二分叉,又形成树枝状的结构,“树枝体”(Dendroid)呈粗短的棒状,长度可达200μm。附枝菌横截面表现为实心的圆球,由暗色泥晶构成,直径较基座菌大,多在50~80μm之间,可将其归属于WOO[37]等划分的类型Ⅰ附枝菌。附枝菌由树形石质均一石中的特征性的凝块状体构成,既能生长在暗色致密泥晶中,也能生长在亮晶方解石胶结物中。
但是对于这种特征性的钙化微生物的生物亲和性却一直备受争议,Riding R[8]和Liu等[35]将附枝菌归为存在疑问的钙化蓝细菌;由于附枝菌有着和现代红藻类似的形态特征故而曾被Luchinina V A和Terleev A A[40]解释为藻类的一种。还有的认为附枝菌是藻类生物在生命周期各个阶段的不同表现形式;Chafetz H S等[41]认为附枝菌应该和肾形菌一样是由细菌诱导的沉淀物而不应该归为光合蓝细菌群落;Adachi N等[42]认为附枝菌是钙化后的结壳而很可能不是蓝细菌,不能将其与葛万菌之类的一起归为钙化蓝细菌,但在次年的著作中他改变了他的看法,将附枝菌归为钙化蓝细菌的一类[43];在现代湖泊相的微生物岩中,存在一种较为典型的灌木丛状组构,Laval等[14]基于该种组构的研究并提出结论,认为附枝菌是受到异养型丝状蓝细菌,如颤菌(Oscillatoria)和眉菌(Calothrix)等菌落的影响,在微生物席内发生钙化作用的产物,而并非通常所认识的是在微生物席表面的钙化作用的产物。不过迄今为止,主流认识认为它同现代大型丝状蓝细菌形成的菌落有一定的相似性[43,44],而且研究表明,一方面,地质记录中所出现的附枝菌似乎并不是通常意义上的细菌细胞体,而是一种可以观察到细菌原始形态的微生物沉积构造,该沉积构造受到细菌钙化作用的影响并在其中保存有完整的细菌个体[2];另一方面,同蓝细菌相似,附枝菌也可能一定程度的趋光性,因而也将其归为蓝细菌种类之一[44];笔者在这里也遵循主流认识,将其归为蓝细菌。
3.2.3 葛万菌(Girvanella)
葛万菌(Girvanella)作为一种由一个相互串联着的细胞列和外部包绕的钙化多糖质黏性胶鞘组成的钙化蓝细菌化石[45],是华北地台寒武系微生物岩中最常见的钙化微生物。在高倍镜下观察构成均一石生物丘的暗色致密泥晶基质以及极不规则的泥晶凝块,可见典型的丝状蓝细菌鞘的钙化作用产物(图8a,图8b为图8a中方框区域的放大;图8c,图8d为图8c中方框区域的放大),这些丝状

图8 均一石生物丘中的丝状钙化蓝细菌
Fig.8 Filamentous calcified cyanobacteria in leiolite bioherms
蓝细菌经活体钙化作用后,其鞘残余物的泥晶壁的厚度较均匀,约为2μm,管内部存在亮晶方解石交代现象,常被解释为蓝细菌细胞列(香毛簇)分解不完全的残余物[7-9,14]。菌丝直径较大(15~20μm)且不相互缠绕的特点,代表着张夏组均一石生物丘中的丝状蓝细菌与现代管状或丝状的第谷菌(Tychonema)具有一定的可类比性[46],所以应该将之归属于与葛万菌的化石组合(Girvanella Group)[8]相类似的全新种类,而不同于类似现代织线菌(Plectonema)[8,46]的、呈缠绕状的较为典型葛万菌(Girvanella)化石。
4 讨论
在河北省秦皇岛市寒武系苗岭统张夏组顶部强迫型海退体系域沉积中存在有由微生物碳酸盐岩主导的生物丘,在顶部集中发育了一套微生物碳酸盐沉淀作用的产物,该产物是高能动荡的浅水环境下所生成的数十米厚的由均一石和树形石质均一石主导的微生物礁,对微生物钙化作用的研究来说具有十分重要的意义。
构成微生物礁的致密泥晶中发育着可以类比于现代胶须菌的基座菌群落、附枝菌以及可以类比于现代织线菌的葛万菌类丝状蓝细菌,葛万菌丝状体内微亮晶充填的管状结构可能为蓝细菌细胞列(香毛簇)降解作用的残余物,而外部均匀的泥晶壁可能是由蓝细菌鞘的活体钙化作用产物和受到胞外聚合物质(Extracellular Polymeric Substance,EPS)的降解过程影响而诱导产生的碳酸盐原地沉淀作用的产物共同组成,蓝细菌鞘的活体钙化作用与大气圈的二氧化碳浓缩机制(CO2 Concentration Mechanisms,CCMs)直接相联系,二氧化碳浓缩机制(CCMs)可以通过调节蓝细菌与外部二氧化碳的交换速率,从而影响环境碱度和碳酸钙的沉淀速率。但是并非所有的蓝细菌都能通过以上过程钙化形成坚硬的岩石,所以不能简单地将张夏组均一石生物丘的形成归因为这些蓝细菌的直接建造物,而且均一石内又普遍发育有窗格状组构,该组构是光合作用过程产生的氧气气泡残余物[33],则更应该将它们解释为有机矿化[47]过程和早期石化作用[1]的产物,这是一个发生在微生物膜或微生物席内的由蓝细菌所主导的复杂微生物调节过程。暗色泥晶凝块间充填的底栖鲕粒,代表着富营养条件下放射显微方解石的微生物沉淀[32],就像梅冥相等[13]所描述的辽东半岛芙蓉统凝块石生物丘中的底栖鲕粒,加之葛万菌之类的丝状体内的微亮晶方解石充填物明显代表着华北地台寒武纪“方解石海”[48]的基本特点,所以在“方解石海”[48]的基本环境下,二氧化硅或者硅酸盐转化为碳酸钙沉淀物的条件是不存在的。
发育在致密泥晶基质和生物潜穴内的白云石晶体,可能与高有机质含量的成岩微环境中的白云石沉淀作用[27]以及选择性白云石化作用相关,这种白云石化机制常发生在高岭石和蒙脱石族矿物中,可能与Mg粘土相关联,其中三八面体硅镁石最具代表性[49]。在鲕粒中的矿化作用主要发生在由纳米纤维构成的微生物席[50]中,该微生物席主要由纤维状的坡缕石(palygorskite)和海泡石(sepiolite)组成,且硅镁石、非晶质镁硅酸盐也存在类似的过程。这一时期,EPS的降解过程会导致释放Mg2+,Mg2+浓度的升高则加速了硫酸盐还原反应的进行,碳酸钙矿物的沉淀则逐渐增多,致使Mg2+浓度升高,Ca2+浓度降低,整体上使Mg2+/Ca2+比值相对升高,从而使发生钙化至成岩作用期间的微生物席或微生物膜发生白云石化,最终可能形成燧石或者一种独特的成岩白云石残余物,代表着一种特别的白云石形成机制而并非GREGG J M等[51]提出的“微生物白云岩”模式。另外致密泥晶基质中还可以观察到黄铁矿晶体残余物,在暗色泥晶中具有更高密度的分布,表明了在富含有机质、呈还原态的大环境中,由硫酸盐还原细菌引起的还原反应是了解微生物沉淀作用和早期石化作用的重要窗口,尤其是该过程中在微生物席内起作用的“碱度发动机”(Alkanility engine)和细胞外聚合物(EPS)两个要素所组成的有机基质,更是生物有机矿化的核心影响因素。二者在碳酸盐矿物的原地沉淀过程中紧密耦合[52],加速微生物新陈代谢作用及速率,使内部碱度增加,同时影响外界环境,使外部碱度增加,从而使碳酸盐的沉淀速率大幅增加[1],形成了秦皇岛苗岭统张夏组顶部的由致密泥晶构成的均一石生物丘,而其微观组构中则存在多样化形态的钙化蓝细菌化石,是了解均一石形成过程中复杂机理和生长样式的关键窗口。
5 结论
均一石生物丘发育在河北秦皇岛寒武系苗岭统张夏组的淹没不整合三级层序的强迫海退体系域沉积的顶部,代表高能浅水环境下,因海平面缓慢下降而发生的碳酸盐沉积。均一石生物丘中发育多样化的钙化蓝细菌,表明这些生物丘形成与蓝细菌所主导的光合微生物膜或微生物席内的复杂钙化作用有着密切的关系。对于许多细节问题仍需要更进一步的研究才能得出合理的解释,但我们针对寒武纪光合微生物席主导的碳酸盐微生物丘的研究能够为日后进一步的研究提供重要的思考途径以及有用的信息。
利益冲突: 作者声明无利益冲突。
[①] *通讯作者 Corresponding author:郭翰良,1041016327@qq.com
收稿日期:2023-07-02; 录用日期:2023-07-14; 发表日期:2023-09-28
基金项目:本项研究得到了国家自然科学基金项目“华北克拉通北缘寒武纪生物丘沉积组构多样性研究”(资助号41492090)的资助
参考文献(References)
[1] Dupraz C, Reid R P, Visscher P T, et al. Microbialites, modern[J]. Encyclopedia of geobiology, 2011: 617-635.
https://doi.org/10.1007/springerreference_187365
[2] 肖恩照, 王皓, 覃英伦. 寒武纪芙蓉统均一石沉积组构及环境特征—以河北涞源长山组为例[J]. 沉积学报, 2020, 38(1): 76-90.
http://dx.doi.org/10.14027/j.issn.1000-0550.2019.025
[3] Braga J C, Martin J M, Riding R. Controls on microbial dome fabric development along a carbonate-siliciclastic shelf-basin transect, Miocene, SE Spain[J]. Palaios, 1995: 347-361.
https://doi.org/10.2307/3515160
[4] 梅冥相. 蓝细菌繁荣滋养的苗岭世光养碳酸盐岩工厂: 以安徽寿县卧龙山剖面崮山组为例[J]. 地质学报, 2021, 95(12): 3571-3591.
http://dx.doi.org/10.3969/j.issn.0001-5717.2021.12.001
[5] 梅冥相. 寒武纪苗岭世特别的光养碳酸盐岩工厂: 以江苏徐州贾旺剖面张夏组为例[J]. 地质学报, 2022, 96(3): 744-768.
http://dx.doi.org/10.3969/j.issn.0001-5717.2022.03.002
[6] Burne R V, Moore L S. Microbialites: organosedimentary deposits of benthic microbial communities[J]. Palaios, 1987: 241-254.
https://doi.org/10.2307/3514674
[7] Riding R. Microbial carbonates: the geological record of calcified bacterial-algal mats and biofilms[J]. Sedimentology, 2000, 47: 179-214.
https://doi.org/10.1046/j.1365-3091.2000.00003.x
[8] Riding R. Cambrian calcareous cyanobacteria and algae [M]//Calcareous algae and stromatolites. Berlin, Heidelberg: Springer Berlin Heidelberg, 1991: 305-334.
https://doi.org/10.1007/978-3-642-52335-9_16
[9] Riding R. Microbialites, stromatolites, and thrombolites [J]. Encyclopedia of geobiology, 2011: 635-654.
https://doi.org/10.1007/springerreference_187366
[10] Riding R. Structure and composition of organic reefs and carbonate mud mounds: concepts and categories [J]. Earth-Science Reviews, 2002, 58(1-2): 163-231.
https://doi.org/10.1016/s0012-8252(01)00089-7
[11] Latif K, Xiao E, Riaz M, et al. Calcified cyanobacteria fossils from the leiolitic bioherm in the Furongian Changshan Formation, Datong(North China Platform)[J]. Carbonates and Evaporites, 2019, 34: 825-843.
https://doi.org/10.1007/s13146-018-0472-8
[12] 梅冥相, MUHAMMAD Riaz, 刘丽, 等. 蓝细菌微生物席主导的芙蓉统均一石生物丘: 以河北涞源祁家峪剖面为例[J]. 地质论评, 2019, 65(6): 1103-1133.
http://dx.doi.org/10.16509/j.georeview.2019.05.004
[13] 梅冥相, 刘丽, 孟庆芬. 光合作用生物膜建造的凝块: 来自于辽东半岛芙蓉统长山组凝块石生物丘中的一些证据[J]. 古地理学报, 2019(2): 254-277.
http://dx.doi.org/10.7605/gdlxb.2019.02.015
[14] 梅冥相, Latif K, 孟庆芬. 寒武系张夏组鲕粒滩中微生物碳酸盐岩主导的生物丘: 以河北秦皇岛驻操营剖面为例[J]. 地质学报, 2019, 93(1): 227-251.
http://dx.doi.org/10.3969/j.issn.0001-5717.2019.01.014
[15] Pruss S B, Finnegan S, Fischer W W, et al. Carbonates in skeleton-poor seas: new insights from Cambrian and Ordovician strata of Laurentia[J]. Palaios, 2010, 25(2): 73-84.
https://doi.org/10.2110/palo.2009.p09-101r
[16] 史晓颖, 陈建强, 梅仕龙. 华北地台东部寒武系层序地层年代格架[J]. 地学前缘, 1997(102): 161-173.
[17] 李晓波, 偶奇, 王旖旎, 等. 辽宁兴城地区前寒武纪地层序列和不整合———兼讨论燕山裂陷槽东南部的沉积古地理演化[J]. 沉积学报, 2020, 38(4): 687-711.
https://doi.org/10.14027/j.issn.1000-0550.2020.004
[18] 潘桂棠, 陆松年, 肖庆辉, 等. 中国大地构造阶段划分和演化[J]. 地学前缘, 2016, 23(6): 1-23.
http://dx.doi.org/10.13745/j.esf.2016.06.001
[19] 梅冥相, MUHAMMAD Riaz, 刘丽, 等. 辽东半岛复州湾剖面寒武系第二统光合作用生物膜建造的核形石[J]. 古地理学报, 2019, 21(1): 31-48.
http://dx.doi.org/10.7605/gdlxb.2019.01.002
[20] 梅冥相, MUHAMMAD Riaz, 孟庆芬, 等. 鲕粒滩相灰岩特别的核形石灰岩帽———以山西繁峙茶坊子剖面寒武系张夏组为例[J]. 地质论评, 2019, 65(04): 839-856.
http://dx.doi.org/10.16509/j.georeview.2019.04.005
[21] 朱一丹, 秦仁月. 河北秦皇岛驻操营剖面张夏组鲕粒滩叠层石生物丘特征及沉积环境[J]. 东北石油大学学报, 2022, 46(2): 23-32.
http://dx.doi.org/10.3969/j.issn.2095-4107.2022.02.003
[22] 梅冥相, 刘丽, 胡媛. 北京西郊寒武系凤山组叠层石生物层[J]. 地质学报, 2015, 89(2): 440-460.
http://dx.doi.org/10.19762/j.cnki.dizhixuebao.2015.02.018
[23] 梅冥相, 张瑞, 李屹尧, 等. 华北地台东北缘寒武系芙蓉统叠层石生物丘中的钙化蓝细菌[J]. 岩石学报, 2017, 33(4): 1073-1093.
[24] 梅冥相. 从正常海退与强迫型海退的辨别进行层序界面对比: 层序地层学进展之一[J]. 古地理学报, 2010, 12(5): 549-564.
http://dx.doi.org/
doi:10.7605/gdlxb.2010.05.005
[25] 齐永安, 孙晓芳, 代明月, 等. 豫西鲁山寒武系馒头组微生物岩旋回及其演化[J]. 微体古生物学报, 2017, 34(2): 170-178.
http://dx.doi.org/10.16087/j.cnki.1000-0674.2017.02.005
[26] Gingras M K, Pemberton S G, Muelenbachs K, et al. Conceptual models for burrow-related, selective dolomitization with textural and isotopic evidence from the Tyndall Stone, Canada[J]. Geobiology, 2004, 2(1): 21-30.
https://doi.org/10.1306/8626de55-173b-11d7-8645000102c1865d
[27] Roberts J A, Kenward P A, Fowle D A, et al. Surface chemistry allows for abiotic precipitation of dolomite at low temperature[J]. Proceedings of the National Academy of Sciences, 2013, 110(36): 14540-14545.
https://doi.org/10.1073/pnas.1305403110
[28] Hunt D, Tucker M E. Stranded parasequences and the forced regressive wedge systems tract: deposition during base-levelfall[J]. Sedimentary Geology, 1992, 81(1-2): 1-9.
https://doi.org/10.1016/0037-0738(92)90052-s
[29] Meng X, Ge M, Tucker M E. Sequence Sequence stratigraphy, sea-level changes and depositional systems in the Cambro-Ordovician of the North China carbonate platform [J]. Sedimentary Geology, 1997, 114(1-4): 189-222.
https://doi.org/10.1306/m57579c1
[30] 倪胜利. 北京西郊下苇甸剖面寒武系叠层石中的底栖鲕粒: 基本特征和重要意义[J]. 地质通报, 2017, 36(2- 3): 485-491.
http://dx.doi.org/10.3969/j.issn.1671-2552.2017.02.030
[31] Gerdes G. What are microbial mats? [J]. Microbial mats: Modern and ancient microorganisms in stratified systems, 2010: 3-25.
https://doi.org/10.1007/978-90-481-3799-2_1
[32] Richter D K, Neuser R D, Schreuer J, et al. Radiaxial-fibrous calcites: A new look at an old problem[J]. Sedimentary Geology, 2011, 239(1-2): 23-36.
https://doi.org/10.1016/j.sedgeo.2011.06.003
[33] Bosak T, Bush J W M, Flynn M R, et al. Formation and stability of oxygen-rich bubbles that shape photosynthetic mats[J]. Geobiology, 2010, 8(1): 45-55.
https://doi.org/10.1111/j.1472-4669.2009.00227.x
[34] 魏柳斌, 赵俊兴, 苏中堂, 等. 鄂尔多斯盆地奥陶系中组合微生物碳酸盐岩分布规律及沉积模式[J]. 石油勘探 与开发, 2021, 48(6): 1162-1174.
http://dx.doi.org/10.11698/PED.2021.06.08
[35] Liu L, Wu Y, Yang H, et al. Ordovician calcified cyanobacteria and associated microfossils from the Tarim Basin, Northwest China: systematics and significance [J]. Journal of Systematic Palaeontology, 2016, 14(3): 183-210.
https://doi.org/10.1080/14772019.2015.1030128
[36] Whitton B A, Mateo P. Rivulariaceae[J]. Ecology of Cyanobacteria II: their diversity in space and time, 2012: 561-591.
https://doi.org/10.1007/978-94-007-3855-3_22
[37] Woo J, Chough S K, Han Z. Chambers of Epiphyton thalli in microbial buildups, Zhangxia formation(Middle Cambrian), Shandong province, China[J]. Palaios, 2008, 23(1): 55-64.
https://doi.org/10.2110/palo.2006.p06-103r
[38] Woo J, Chough S K. Growth patterns of the Cambrian microbialite: phototropism and speciation of Epiphyton [J]. Sedimentary Geology, 2010, 229(1-2): 1-8.
https://doi.org/10.1016/j.sedgeo.2010.05.006
[39] 殷振轩, 左银辉, 肖恩照. 华北地台寒武系芙蓉统均一石沉积组构分析[J]. 东北石油大学学报, 2022, 46(3): 45-53, 65.
http://dx.doi.org/10.3969/j.issn.2095-4107.2022.03.004
[40] Luchinina V A. Remalcis and Epiphyton as different stages in the life cycle of calcareous algae[J]. Paleontological Journal, 2009, 43: 463-468.
https://doi.org/10.1134/s0031030109040169
[41] Chafetz H S, Guidry S A. Bacterial shrubs, crystal shrubs, and ray-crystal shrubs: bacterial vs. abiotic precipitation[J]. Sedimentary Geology, 1999, 126(1- 4): 57-74.
https://doi.org/10.1016/s0037-0738(99)00032-9
[42] Adachi N, Nakai T, Ezaki Y, et al. Late Early Cambrianarchaeocyath reefs in Hubei Province, South China: modes of construction during their period of demise[J]. Facies, 2014, 60: 703-717.
https://doi.org/10.1007/s10347-013-0376-y
[43] Adachi N, Kotani A, Ezaki Y, et al. Cambrian Series 3lithistid sponge-microbial reefs in Shandong Province, North China: reef development after the disappearanceof archaeocyaths[J]. Lethaia, 2015, 48(3): 405-416.
https://doi.org/10.1111/let.12118
[44] 韩作振, 陈吉涛, 张晓蕾, 等. 鲁西寒武系第三统张夏组附枝菌与附枝菌微生物灰岩特征研究[J]. 地质学报, 2009, 83(8): 1097-1103.
http://dx.doi.org/10.3321/j.issn:0001-5717.2009.08.006
[45] 齐永安, 张喜洋, 代明月, 等. 豫西寒武系微生物岩中的葛万菌化石及其微观结构[J]. 古生物学报, 2017, 36(2): 154-167.
http://dx.doi.org/10.19800/j.cnki.aps.2017.02.003
[46] de los Ríos A, Ascaso C, Wierzchos J, et al. Microstructureand cyanobacterial composition of microbial matsfrom the High Arctic[J]. Biodiversity and Conservation, 2015, 24: 841-863.
https://doi.org/10.1007/s10531-015-0907-7
[47] Dupraz C, Reid R P, Braissant O, et al. Processes of carbonateprecipitation in modern microbial mats[J]. Earth-Science Reviews, 2009, 96(3): 141-162.
https://doi.org/10.1016/j.earscirev.2008.10.005
[48] Kiessling W. Geologic and biologic controls on the evolutionof reefs[J]. Annual Review of Ecology, Evolution, and Systematics, 2009, 40: 173-192.
https://doi.org/10.1146/annurev.ecolsys.110308.120251
[49] Burne R V, Moore L S, Christy A G, et al. Stevensite inthe modern thrombolites of Lake Clifton, Western Australia: A missing link in microbialite mineralization?[J]. Geology, 2014, 42(7): 575-578.
https://doi.org/10.1130/g35484.1
[50] Perri E, Tucker M E, Słowakiewicz M, et al. Carbonateand silicate biomineralization in a hypersaline microbialmat(Mesaieed sabkha, Qatar): roles of bacteria, extracellularpolymeric substances and viruses[J]. Sedimentology, 2018, 65(4): 1213-1245.
https://doi.org/10.1111/sed.12419
[51] Gregg J M, Bish D L, Kaczmarek S E, et al. Mineralogy, nucleation and growth of dolomite in the laboratoryand sedimentary environment: a review[J]. Sedimentology, 2015, 62(6): 1749-1769.
https://doi.org/10.1111/sed.12202
[52] 辛浩, 肖恩照, 覃英伦, 等. 鲁西地区仙人洞剖面凤山组大型柱状叠层石沉积特征[J]. 东北石油大学学报, 2019, 43(3): 1-11.
Microbial Reefs Predominated by Leiolites in the Zhanxia Formation of Cambrian from Qinhuangdao City,Hebei Province
GUO Hanliang1,*, QIN Renyue2
(1. School of Earth Science and Resources, China University of Geosciences, Beijing 100083, China
2. Guizhou Wu River Energy Investment Co. , Ltd. Guizhou Shale gas Exploration and Development Co. , Ltd, Zunyi 563400, China)
Abstract: Stromatolites, thrombolites, leiolites and dendrites are juxtaposed and classified as typical microbial carbonates, with leiolite characterized by cryptocrystalline lithology, undeveloped agglomerate structure, lacking obvious macroscopic structure and clot. Since its designation in 1995, leiolites have been rarely observed and recognized in strata and modern analogies are scarce, whoch makes the report of a leiolite example extremely important. In order to research the characteristics of leiolites in Cambrian of North China Platform, combine the methods of field sampling observation and observation of indoor microscopic characteristics under the microscope and selected microbial reefs predominated by leiolites with thickness of tens of meters develop intensively at the top of the Forced Regression system domain of the Cambrian Zhangxia Formation in Qinhuangdao, Hebei Province was selected, which constitutes a three-order sequence as research object. Leiolite consists of dense mud crystals and a small amount of microlithophores, and the Filamental Girvanella similar to the modern Plectonema is visible in the dense mud crystals that make up leiolites, while Epiphyton and Hedstroemia can be seen locally in the bushy mud crystals. It is indicated that microbial reefs predominated by leiolites are influenced by complex organic mineralization processes or by early cyanobacteria-dominated lithification occurring within microbial membranes or microbial mats. Therefore, in understanding the formation of ancient microbial reefs, the microbial reef predominated by leiolites at the Cambrian Zhangxia Formation in Qinhuangdao, formed by the calcification of microbial mats, are an important window.
Keywords: Qinhuangdao city, Zhangxia Formation, lieolite, microbial carbonate, calcified cyanobacteri
DOI: 10.48014/cesr.20230702002
Citation: GUO Hanliang, QIN Renyue. Microbial reefs predominated by leiolites in the Zhanxia Formation of Cambrian from Qinhuangdao city, Hebei Province[J]. Chinese Earth Sciences Review, 2023, 2(3): 19-31.
