华南早奥陶世微生物礁中的凝块石:以宜昌黄花场剖面为例
唐浩博
(中国地质大学 (北京) 地球科学与资源学院, 北京 100083)
摘要: 湖北宜昌黄花场剖面红花园组发育一套微生物礁, 微生物礁中保存有良好的微生物碳酸盐岩 (即凝块石) 。基于前人对华南早奥陶世生物礁中后生动物和后生动物群落造礁作用的研究, 本文通过野外实地考察及采样、室内磨片观察, 结合微生物礁的宏观、微观特征, 主要研究微生物礁中凝块石的形成机制。结果发现: 黄花场剖面红花园组微生物礁以凝块石为主, 伴随少量瓶筐石、石海绵、苔藓虫等后生动物化石, 代表了早奥陶世一种特殊的微生物礁类型, 即微生物礁以微生物碳酸盐岩为主, 局部有后生动物参与造礁, 是生物礁类型由微生物礁向后生动物礁过渡过程中一种特殊的生物礁类型。同时, 凝块石的内部发现有大量的钙化蓝细菌化石, 如葛万菌、束线菌及努亚藻, 较为直观的钙化蓝细菌表明了凝块石是由蓝细菌主导的微生物膜或微生物席的沉淀、粘聚及早期石化作用的产物, 表明了凝块石与蓝细菌主导的微生物席复杂钙化作用之间有着十分密切的关系。本文为华南早奥陶世微生物礁中凝块石的形成机制及微生物礁向后生动物礁转变的过渡时期的研究提供了一个较为典型的实例。
关键词: 早奥陶世, 微生物礁, 凝块石, 钙化蓝细菌化石, 黄花场剖面, 华南板块
DOI: 10.48014/cesr.20241011001
引用格式: 唐浩博. 华南早奥陶世微生物礁中的凝块石: 以宜昌黄花场剖面为例[J]. 中国地球科学评论, 2024, 3(4): 196-206.
文章类型: 研究性论文
收稿日期: 2024-10-11
接收日期: 2024-10-21
出版日期: 2024-12-28
0 引言
凝块石作为微生物碳酸盐岩的一种[1-3],最早是由AITKEN[4]在研究加拿大南部落基山脉的寒武纪-奥陶纪特殊的沉积岩时所提出,最初是指“与叠层石相关的,但宏观上以凝块状组构而区别于叠层石纹层组构的隐藻构造”。之后,SHAPIRO[5]将凝块石定义为“由微凝块或中凝块组成的微生物岩”。奥陶纪是生物礁演化历史上一个非常独特的时期,它见证了生物礁组构的复杂性变化,即早奥陶世以微生物礁为主,局部发育有后生动物礁[6-8],之后随着后生动物层孔虫、苔藓虫和珊瑚的蓬勃发展和扩张,中-晚奥陶世则转变为珊瑚-层孔虫-苔藓虫后生动物礁为主[9]。
早奥陶世生物礁在我国华南、华北及塔里木板块均有着广泛分布[10],前人对华南板块早奥陶世生物礁进行了较为详细的研究,如朱忠德等[10]从生物礁的造礁生物、含礁层系的沉积相、生物礁的特征及演化和生物礁的含油性等方面对华南中扬子台地早奥陶世生物礁进行了详细研究;ADACHI等[11]通过对扬子板块边缘和中心发育的生物礁进行对比、归纳和总结后发现,后生动物主导的生物礁在三峡及临近区域最早出现,华南板块三峡地区是研究微生物礁向后生动物生物礁转变最为有利的地区之一;ZHANG等[12]通过对华南板块上、中、下扬子台地寒武纪-早奥陶世发育的生物礁进行对比、研究后,总结归纳出全球气候变化是生物礁演化的主控因素。
湖北省宜昌市位于华南板块的三峡地区,其下奥陶统出露完整、露头情况良好,是研究微生物礁向后生动物生物礁转变最为有利的地区之一。黄花场剖面下奥陶统红花园组中发育一套微生物礁。该微生物礁中保存有良好的微生物碳酸盐岩凝块石。本文在前人对华南早奥陶世生物礁中后生动物或后生动物群落造礁作用研究的基础上,通过野外实地考察及采样、室内磨片观察,详细描述了黄花场剖面红花园组微生物礁的宏观、微观特征,旨在:(1)对黄花场剖面红花园组微生物礁中的造礁组分进行研究;(2)对微生物礁中凝块石的形成机制进行研究分析。本文为华南早奥陶世微生物礁中凝块石的形成机制及微生物礁向后生动物礁转变的过渡时期研究提供了一个较为典型的实例。
1 地质背景
早奥陶世,华南板块位于南半球中低纬度地区,是冈瓦纳古陆西北缘的一部分[13]。此时的华南板块地处低纬度温暖气候区,底栖海洋生物大量繁盛,形成了大量的生物礁[14]。华南板块主要由扬子地块和华夏地块组成[15],其中扬子地块在经历了早奥陶世特马豆克期的海进之后,除了西缘、西北缘仍保留有古陆,其余地方均被海水淹没[15]。因此,扬子地块沿西北向东南方向依次发育了潮坪-潟湖、开阔台地、台地边缘浅滩三个沉积相带(图1a)。此时,除了西部、西北部近岸带有少量陆源沉积物输入以外,在远岸区形成了大量浅海碳酸盐沉积物[16]。
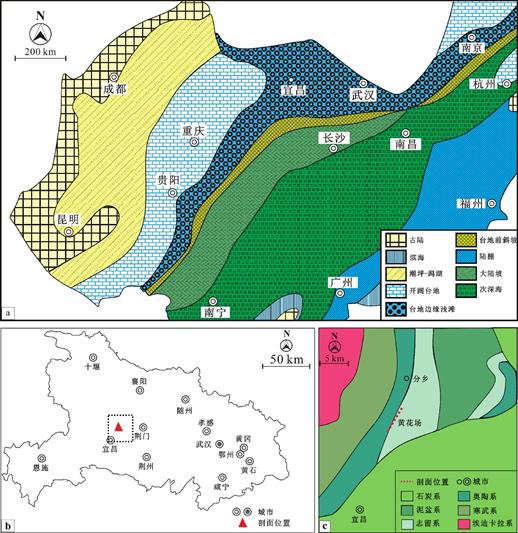
图1 湖北宜昌黄花场剖面的剖面位置
Fig.1 The location of the Huanghuachang section in the Yichang,Hubei province
黄花场剖面位于湖北省宜昌市东北方向23km外的黄花场村(30°51'33″N,111°22'28″E),(图1b和图1c,图1b中黑色线框的放大),是全球中-下奥陶统底界(大坪阶)的界线层型剖面和点位(GSSP)[17]。在早-中奥陶世时,黄花场地区位于扬子碳酸盐台地的中北部,属于台地边缘浅滩沉积相带(图1a),发育了大片以蓝细菌为主的粘结灰岩及生物礁灰岩。同时,黄花场剖面的早奥陶世地层出露连续、完整,含有丰富的海绵、腕足、头足等动物化石。红花园组在时代上属于弗洛期早期沉积,广泛分布于整个扬子地块[18],岩性主要以生物碎屑灰岩、亮晶灰岩等为主[18]。
在通往黄花场剖面的道路边上,早-中奥陶世地层出露良好。沿着道路追索后发现,早-中奥陶世叠层由下至上为南津关组、分乡组、红花园组、大湾组,地层倾角约10°~12°。其中红花园组厚度约20m,其底部发育的生屑灰岩整合覆盖在分乡组(对应于特马豆克阶上部)页岩主导的地层之上(图2a),代表了相对海平面快速下降阶段的产物。顶部则是被大湾组泥晶灰岩所覆盖(图2b和图2c,图2b中白色线框的放大)。红花园组除发育大套生屑灰岩以外,局部仍发育薄层似球粒灰岩及少量页岩,向上生屑灰岩和似球粒灰岩层厚逐渐变薄。

图2 黄花场剖面下奥陶统红花园组岩性柱状图(修改自参考文献[8])
Fig.2 Lithologic column chart of the lower Ordovician Honghuayuan Formation at Huanghuachang section(Modified from reference [8])
2 红花园组微生物礁的宏观特征
黄花场剖面红花园组的微生物礁,宽约4m,高约1~2.5m,呈中间稍高两端略低的丘状产出(图3a和图3b,图3a中白色线框的放大)。野外观察发现,生物礁主要由凝块石构成(图3b,图4a,图4b),礁中发育少量的后生动物如瓶筐石、石海绵等(图4c和图4d)。凝块石的内部组构主要是由暗色致密泥晶组成的、非纹层状的微凝块构成(图4a和图4b)。这些微凝块大都比较小,约1 mm,但也可观察到一些较大的复合型凝块,这些微凝块多呈现不规则状,分布较为均匀,彼此之间很少接触,可以将其归为网格状凝块石的一种。微凝块之间主要由白色致密的泥晶方解石充填(图4a和图4b)。微生物礁中发育的后生动物化石主要以瓶筐石为主(图4c和图4d),瓶筐石轮廓清晰、出露良好,大小约1cm(图4c)。在垂直于瓶筐石的表面,可以观察到其清晰的椭圆形轮廓,同时也可以观察到瓶筐石明显的双层体壁(图4c和图4d),外体壁与内体壁间距离约0.4cm,风化后的瓶筐石中腔明显高于双壁(图4c),相反,新鲜面上则与中腔平齐(图4d)。在生物礁表面还可以观察到可能的石海绵化石(图4c),石海绵相比于瓶筐石较小,直径约0.5cm,有着类似于瓶筐石的双层体壁,内、外壁间约0.2cm,两者均为海绵动物的一种。

图3 黄花场剖面红花园组微生物礁的宏观特征
Fig.3 Macroscopic characterization of microbial reef in the Honghuayuan Formation at the Huanghuachang section

图4 黄花场剖面红花园组微生物礁的宏观特征
Fig.4 Macroscopic characterization of microbial reef in the Honghuayuan Formation at the Huanghuachang section
3 红花园组微生物礁的微观特征
3.1 整体微观特征
凝块石在显微镜下由大小各异的凝块及凝块间充填的亮晶方解石构成(图5a和图5b)。这些大小不同的、形态不规则的凝块主要由暗色致密泥晶组成,大小多在0.2~1mm之间,属于微凝块的范畴,但也有超过1mm的复合型凝块-葡萄石的发育(图5a和图5b),这种复合型凝块由致密的暗色泥晶包裹几个微凝块组成,在致密泥晶中还可见三叶虫骨骼碎片等生物碎屑化石(图5c和图5d)。这些在低倍显微镜下呈现暗色致密泥晶的微凝块,在高倍显微镜下,可以观察到高密度保存的钙化蓝细菌化石。这些钙化蓝细菌化石主要有互相缠绕的可以类比于现代织线菌的葛万菌(Girvanella)化石(图6a,图6b,图6e)、可以类比于现代的微鞘菌[19]或鞘线菌的束线菌(Subtifloria)化石(图6c和图6d)及一种疑难钙化微体化石-努亚藻(Nuia)(图6f)。除了钙化蓝细菌化石以外,还可以观察到清楚的瓶筐石(图5c,红色箭头)、石海绵(图5c和图5d,白色箭头)、苔藓虫(图5e,红色箭头)等多样化的后生动物化石。
3.2 微生物礁中后生动物化石的微观特征
微生物礁中后生动物的数量虽然占比较小,但仍发育早奥陶世后生动物组合的代表生物,如瓶筐石(图5c,红色箭头)、石海绵(图5c和图5d,白色箭头)及苔藓虫(图5e,白色箭头)。前人对华南早奥陶世的生物礁进行了详细的研究后[20],发现生物礁中都发育着保存程度不同的石海绵。在显微镜下,石海绵形状主要为片状,宽约2~4mm,高约3mm,保存为具有明显轮廓的致密泥晶中的海绵骨针,未见完整的石海绵化石。这些石海绵常与瓶筐石相伴随出现,同时在包裹海绵骨针的致密泥晶中还可观察到小的底栖鲕粒和生物碎屑(图5e)。
瓶筐石在显微镜下由内、外两层体壁组成,内壁厚度较小,约200μm,外壁相对较厚,约500μm,个体宽约4mm(图5c),在瓶筐石壁外构造下可隐约见到葛万菌化石(图5e和图5f),可能是发育葛万菌的微生物膜附着在瓶筐石壁外构造之上,通过微生物膜的粘附性,加固微生物礁[7]。在显微镜下,还可以观察到一个完整的苔藓虫,宽约0.5mm,长约3mm,发育在致密泥晶之中,附着在石海绵之上(图5d),局部形成一种苔藓虫-石海绵组合,即苔藓虫通过附着在石海绵之上,局部加固微生物礁的稳定性[7]。瓶筐石壁间充填有三叶虫、腕足等生物碎屑和致密泥晶[21]。
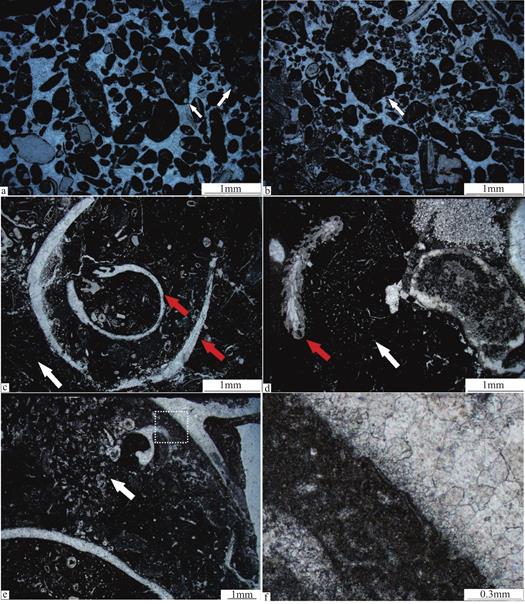
图5 红花园组微生物礁中后生动物化石的微观特征
Fig.5 Microscopic characteristics of metazoan fossils in microbial reefs of the Honghuayuan Formation
3.3 微生物礁中钙化蓝细菌化石的微观特征
在组成凝块石的微凝块中,可以观察到保存精美的葛万菌化石,其通常由两部分组成,一部分是泥晶或微亮晶方解石构成的管状或丝状体,另一部分则是包裹管状或丝状体的较薄泥晶壁,化石彼此间高度缠绕(图6a,图6b(图6a中白色线框的放大),图6e)。葛万菌化石的泥晶壁厚度通常不大,为1~3μm,泥晶或微亮晶方解石构成的管状或丝状体宽10μm左右,管状或丝状体的横切面为内亮外暗的圆形构造。这种被暗色泥晶壁所包裹的微亮晶方解石丝可能代表着一种丝状蓝细菌活体钙化作用的残余物[22],内部的微亮晶方解石细丝代表着蓝细菌细胞列降解作用残余物[2,23],这种葛万菌化石可以类比于现代微生物席中发现的织线菌。
在拉长状凝块中,还可以观察到清楚的束线菌(Subtifloria)化石。这一类的丝状蓝细菌化石以较长的丝状体(长度可达0.5 mm,甚至更长)、较薄且厚度均匀的泥晶壁、丝状体直径小于10 μm、呈束状密集生长为特征(图6c,图6d),与高度缠绕的葛万菌化石有着明显的不同。束线菌则可以类比于现代的微鞘菌[19]或鞘线菌。
努亚藻(Nuia)在镜下表现为椭圆形和近圆形,直径约0.1mm(图6f)。主要由中央的黑色区域、放射状方解石显微充填的圈层及最外层的钙化外鞘组成,可能是蓝藻门念珠藻目胶须藻科的早期化石代表[24]。
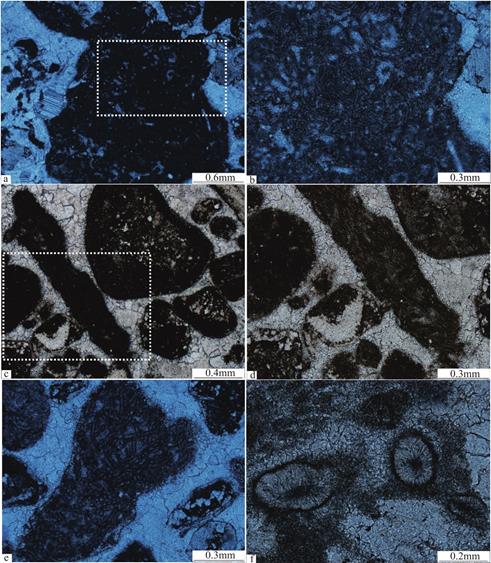
图6 红花园组微生物礁中钙化蓝细菌化石的微观特征
Fig.6 Microscopic characteristics of calcified cyanobacteria fossils in microbial reefs of the Honghuayuan Formation
4 讨论
黄花场剖面微生物礁主要发育微生物碳酸盐岩-凝块石(图4a,图4b,图5a,图5b,图6),少量发育瓶筐石(图4c,图4d,图5c)、石海绵(图5c,图5d)、苔藓虫(图5e)等多样化的后生动物化石。这种既发育微生物碳酸盐岩-凝块石,同时又发育少量后生动物化石的微生物礁,既与寒武纪苗岭世-芙蓉世发育的由微生物碳酸盐岩主导的、无后生动物化石的微生物礁/丘有所不同[25-31],又区别于中-晚奥陶世以珊瑚-层孔虫-苔藓虫为主的后生动物生物礁[9]。代表了早奥陶世早期一种较为特殊的微生物礁类型,即:微生物礁的组分仍像寒武纪苗岭世-芙蓉世发育的生物礁一样以微生物碳酸盐岩为主,与之不同的是,微生物礁中局部有着后生动物参与造礁,代表了生物礁由微生物礁向后生动物礁转变的一个过渡类型。
微生物礁中的凝块石在镜下由典型的凝块或者微凝块组成(图6),符合凝块石由“由微凝块(中凝块)组成的微生物岩”的定义,是一种较为典型的凝块石[5,32,33]。关于这种较为典型的凝块石的形成机制,前人已经做出了很多的研究工作,认为凝块石主要的形成机制有:(1)凝块石是由叠层石受到后生动物的扰动作用、生物潜穴和钻孔作用或强烈的水动力条件改造所形成,这种成因的凝块石常见于前寒武纪末期及古生代地层中[34,35];(2)凝块石像叠层石一样,由一些原生微生物的钙化作用和粘聚作用形成,呈柱状、丘状和层状产于潮下环境[1,36];(3)可能代表了一种真核生物构造[37]。
通过对红花园组微生物礁中凝块石的宏观、微观特征研究后发现,在凝块石中并未发现有明显的生物扰动、生物潜穴及钻孔现象,表明红花园组微生物礁中发育的凝块石并非是由叠层石受到后生动物的扰动作用、生物潜穴和钻孔作用或强烈的水动力条件改造所形成。在凝块石中,也未发育有真核生物,故红花园组微生物礁中的凝块石也并非代表了一种真核生物构造。同时,红花园组微生物礁中凝块石的微观特征表明,凝块石主要由暗色泥晶组成的凝块和凝块间的亮晶方解石构成。在低倍显微镜下呈现的暗色致密泥晶的凝块,在高倍显微镜下可以观察到丰富的葛万菌(Girvanella)、束线菌(Subtifloria)、努亚藻(Nuia)等钙化蓝细菌化石。在一些凝块的边缘还可以见到可能的黄铁矿晶体的残余物(图6(f)),这种黄铁矿晶体的残余物可能是由硫酸盐还原细菌所主导的硫酸盐还原反应形成[38]。凝块中发育的多样化且保存精美的蓝细菌化石及与硫酸盐还原细菌有关的黄铁矿晶体,说明了凝块石正是由多样化的钙化蓝细菌组成的微生物膜或微生物席的沉淀、粘聚及早期石化作用的产物,表明了凝块石与蓝细菌主导的微生物席复杂钙化作用之间存在有十分密切的关系。
凝块石中的凝块形成主要包括两种形式的钙化作用:(1)蓝细菌自身的钙化作用:凝块石中发育丰富的丝状钙化蓝细菌及可能的黄铁矿晶体的残余物,表明蓝细菌所进行的光合作用及硫酸盐还原菌主导的硫酸盐还原反应可以提高周围环境的碱度,从而促进CaCO3沉淀,使得CaCO3过饱和,最终导致碳酸盐矿物发生沉积和早期石化的微生物席的形成[1,39];(2)细胞外聚合物质(EPS)的吸附和钙化作用[40]:由蓝细菌分泌的EPS,会从海水中吸附Ca2+。但当这种吸附作用超过EPS自身吸附Ca2+的能力时,EPS就会发生降解,释放出Ca2+、Mg2+、H+,从而使得碳酸盐晶体在微生物膜或微生物席内发生钙化,形成凝块石中的凝块[41]。
5 结论
黄花场剖面红花园组微生物礁以凝块石为主,伴随少量瓶筐石、石海绵、苔藓虫等后生动物化石,代表了早奥陶世一种特殊的微生物礁类型,即微生物礁以微生物碳酸盐岩为主,局部有后生动物参与造礁,是生物礁类型由微生物礁向后生动物礁过渡过程中一种特殊的生物礁类型。同时,凝块石的内部发现有大量的钙化蓝细菌化石,如葛万菌、束线菌及努亚藻,较为直观的钙化蓝细菌表明了凝块石是由蓝细菌主导的微生物膜或微生物席的沉淀、粘聚及早期石化作用的产物,表明了凝块石与蓝细菌主导的微生物席复杂钙化作用之间有着十分密切的关系。这种密切的关系中仍留有许多细节上的问题需要我们进一步的研究才能得到更好的解决,如形成凝块石的钙化蓝细菌化石的精确物种归类、蓝细菌在凝块石形成过程中的代谢作用形式。因此,对于微生物席内的钙化蓝细菌化石的精确物种归类及内部生物颗粒成因可能是未来微生物碳酸盐岩研究过程中所面临的最大挑战。
利益冲突: 作者声明无利益冲突
[④] 通讯作者 Corresponding author:唐浩博,1508499253@qq.com
收稿日期:2024-10-11; 录用日期:2024-10-21; 发表日期:2024-12-28
基金项目:本文得到了国家自然科学基金项目(41492090)“华北克拉通北缘寒武纪生物丘沉积组多样性研究”的资助
参考文献(References)
[1] Riding R. Microbial carbonates: the geological record of calcified bacterial-algal mats and biofilms[J]. Sedimentology, 2000, 47(1): 179-214.
https://doi.org/10.1046/j.1365-3091.2000.00003.x
[2] Riding R. Calcified cyanobacteria [J]. Encyclo-pedia of Earth Sciences Series, 2011, 211-223.
https://doi.org/10.1007/978-1-4020-9212-1_63
[3] Lee J H, Riding R. Stromatolite-rimmed thro-mbolite columns and domes constructed by microstromatolites, calcimicrobes and sponges in late Cambrian biostromes, Texas, USA[J]. Sedimentology, 2023, 70(2): 293-334.
https://doi.org/10.1111/sed.13048
[4] Aitken J D. Classification and environmental significance of cryptalgal limestones and dolomites, with illustrations from the Cambrian and Ordovician of southwestern Alberta[J]. Journal of Sedimentary Research, 1967, 37(4): 1163-1178.
https://doi.org/10.1306/74D7185C-2B21-11D7-8648000102C1865D
[5] Shapiro R S. A Comment on the Systematic Co-nfusion of Thrombolites[J]. Palaios, 2000, 15(2): 166-169.
https://doi.org/10.1669/08831351(2000)015
[6] Adachi N. , Ezaki Y. , Liu J. , et al. Early Ordovician reef construction in Anhui Province, South China: a geobiological transition from microbial-to metazoan-dominant reefs[J]. Sedimentary Geology, 2009, 220(1-2): 1-11.
https://doi.org/10.1016/j.sedgeo.2009.05.012
[7] Adachi N, Ezaki Y, Liu J B. Early Ordovician reef construction shift in reef construction from microbial to metazoan reefs[J]. Palaios, 2011, 26(1-2): 106-114.
https://doi.org/10.2110/palo.2010.p10-097r
[8] Li Q J, Li Y, Wang J P, et al. Early Ordovician lithistid sponge– Calathium reefs on the Yangtze Platform and their paleoceanographic implications[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2015, 425(0): 84-96. 10. 1016/j. palaeo. 2015. 02. 034
[9] Webby B D. Patterns of Ordovician reef development[J]. Special Publications, 2002, 72: 129-179.
https://doi.org/10.2110/pec.02.72.0129
[10] 朱忠德, 胡明毅, 刘秉理, 等. 中国早中奥陶世生物礁研究[M]. 北京: 地质出版社, 2006: 1-43.
[11] Adachi N, Ezaki Y, Liu J B, et al. Cambrian t-hrough Ordovician reef transitions in North and South China: Changes in reef construction and background geobiological environments[J]. Pala-eogeography, Palaeoclimatology, Palaeoecology, 2023, 630: 111804.
https://doi.org/10.1016/j.palaeo.2023.111804
[12] Zhang Y Y, Li Q J, Li Y, et al. Cambrian to Lower Ordovician reefs on the Yangtze Platform, South China Block, and their controlling factors[J]. Facies, 2016, 62(3): 1-18.
https://doi.org/10.1007/s10347-016-0466-8
[13] Torsvik T, Cocks L. New global palaeogeographical reconstructions for the Early Palaeozoic and their generation[J]. Early Palaeozoic Biogeography and Palaeogeography, 2013, 38: 5-24.
https://doi.org/10.1144/M38.2
[14] Wang J P, Li Q J, Kershaw S, et al. Late Tre-madocian(Early Ordovician)reefs on the Yangtze Platform, South China, and their geobiological im-plications: a synthesis[J]. Journal of Palaeogeog-raphy, 2021, 10(2): 182-196.
https://doi.org/10.1186/s42501-021-00086-0
[15] 周名魁, 王汝植, 李志明, 等. 中国南方奥陶-志留纪岩相古地理与成矿作用[M]. 北京: 地质出版社, 1993: 1-115.
[16] ZHAN R B, JIN J S. Ordovician-Early Silurian(Llandovery)Stratigraphy and Palaeontology of t-he Upper Yangtze Platform, South China[M]. Beijing: Science Press, 2007: 1-169.
[17] Wang X F, Stouge S, Erdtmann B. , et al. A proposed GSSP for the base of the Middle Ordovician Series: the Huanghuachang section, Yichang, China[J]. Episodes, 2005, 28(2): 105-117.
https://doi.org/10.18814/epiiugs/2005/v28i2/004
[18] 张元动, 詹仁斌, 袁文伟, 等. 中国奥陶纪岩石地层划分和对比[J]. 地层学杂志, 2021, 45(3): 250-270. 10. 19839/j. cnki. dcxzz. 2021. 0023
[19] Riding R. Classification of Microbial Carbonates[J]. Calcareous Algae and Stromatolites, 1991: 21-51.
https://doi.org/10.1007/978-3-642-52335-9_2
[20] Adachi N, Liu J B, Ezaki Y. Early Ordovician reefs in South China(Chenjiahe section, Hubei Province): deciphering the early evolution of skeletal-dominated reefs[J]. Facies, 2013, 59: 451-466.
https://doi.org/10.1007/s10347-012-0308-2
[21] 党志英, 沈玉林, 杨天洋, 金军, 赵勇. 黔西水城长兴期 晚期生物与环境突变响应: 基于古生物学和地球化学证据[J]. 古地理学报, 2023, 25(6): 1421-1436.
https://doi.org/10.7605/gdlxb.2023.04.049
[22] 肖恩照, 王皓, 覃英伦, 等. 寒武纪芙蓉统均一石沉积组 构及环境特征———以河北涞源长山组为例[J]. 沉积 学报, 2020, 38(1): 76-90.
https://doi.org/10.14027/j.issn.1000-0550.2019.025
[23] 梅冥相, LATIF K, 刘丽, 等. 光合作用生物膜建造的凝块: 来自于辽东半岛芙蓉统长山组凝块石生物丘中的一些证据[J]. 古地理学报, 2019, 21(2): 254-277.
https://doi.org/10.7605.gdlxb.2019.02.015
[24] 王冬梅, 杨宇宁, 刘伟, 等. 黔北早奥陶世疑难钙藻化石Nuia形态分类与钙化机制[J]. 地质学报, 2023, 97(7): 2093-2110.
https://doi.org/10.19762/j.cnki.dizhixuebao.2022050
[25] 王龙, LATIF K, RIAZ M, 等. 微生物碳酸盐岩的成因、分类以及问题与展望———来自华北地台寒武系微生物碳酸盐岩研究的启示[J]. 地球科学进展, 2018, 33(10): 1005-1023.
https://doi.org/10.11867/j.issn.1001-8166.2018.10.1005
[26] 王龙. 太行山中南部及其邻区寒武系微生物碳酸盐岩主导的生物丘研究[D]. 北京: 中国地质大学(北京), 2019.
https://doi.org/10.27493/d.cnki.gzdzy.2019.000145
[27] 秦仁月. 辽东半岛复州湾剖面寒武系馒头组核形石沉积特征[J]. 东北石油大学学报, 2020, 44(5): 46-58, 7-8.
https://doi.org/10.3969/j.issn.2095-4107.2020.05.005
[28] 辛浩, 肖恩照, 王龙, 等. 肥城邓家庄剖面寒武系微生物碳酸盐岩沉积序列[J]. 沉积学报, 2020, 38(1): 64-75.
https://doi.org/10.14027/j.issn.1000-0550.2019.071
[29] 柴凯旗, 秦仁月. 光合微生物膜钙化建造的底栖鲕粒———以辽西葫芦岛三道沟剖面张夏组鲕粒滩为例[J]. 东北石油大学学报, 2021, 45(3): 73-85, 9.
https://doi.org/10.3969/j.issn.2095-4107.2021.03.008
[30] 梅冥相. 光合作用微生物席主导的寒武系苗岭统崮山组均一石:以山东省泗水县圣水峪剖面为例[J]. 古地理学报, 2021, 23(2): 335-358.
https://doi.org/10.7605/gdlxb.2021.02.022
[31] Adachi Natsukawa H, Ezaki Y, et al. Cambrian Series 2(Stage 4)calcimicrobial reefs in Shand-ng Province, North China: Calcimicrobial diver-ity and contributionto reef construction[J]. Pa-aeogeography, Palaeoclimatology, Palaeoecology, 2023, 610: 111351.
https://doi.org/10.1016/j.palaeo.2022.111351
[32] Xin H, Chen J T Gao B, et al. Spatio-temporal distributionof the Cambrian maceriate reefs across the NorthChina Platform[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2023, 614: 111429.
https://doi.org/10.1016/j.palaeo.2023.111429
[33] 吴亚生. 生物岩的分类 [J]. 古地理学报, 2023, 25(3): 511-523.
https://doi.org/10.7605/gdlxb.2023.03.018
[34] 曹瑞骥, 袁训来. 叠层石[M]. 合肥: 中国科学技术大学出版社, 2006: 1-383.
[35] 梅冥相. 从凝块石概念的演变论微生物碳酸盐岩的研究进展[J]. 地质科技情报, 2007, 26(6): 1-9.
[36] 张震武, 肖恩照, 覃英伦, 等. 莱芜雪野剖面寒武系苗岭统凝块石沉积特征[J]. 东北石油大学学报, 2019, 43(3): 67-77, 124.
https://doi.org/10.3969/j.issn.2095-4107.2019.03.007
[37] Feldmann M, Mckenzie J A. Stromatolite-throm-boliteassociations in a modern environment, Le-e StockingIsland, Bahamas [J]. Palaios, 1998, 13(2): 201-212.
https://doi.org/10.2307/3515490
[38] 辛浩, 肖恩照, 覃英伦, 等. 鲁西地区仙人洞剖面凤山组大型柱状叠层石沉积特征[J]. 东北石油大学学报, 2019, 43(3): 1-11.
https://doi.org/10.3969/j.issn.2095-4107.2019.03.001
[39] Dupraz C, Reid R P, Braissant O, et al. Processes of carbonateprecipitation in modern microbial mats[J]. Earth-Science Reviews, 2009, 96(3): 141-162.
https://doi.org/10.1016/j.earscirev.2008.10.005
[40] Decho A. Microbial exopolymer secretions in ocean enviroments-their role(s)in food webs and marine processes[J]. Oceanography and Marine Biology, 1990, 28: 73-153.
[41] Wilmeth D T, Corsetti F A, Bisenic N, et al. Punctuatedgrowth of microbial cones within early cambrian oncoids, bayan gol formation, western Mongolia(Article)[J]. Palaios, 2015, 30(12): 836-845.
https://doi.org/10.2110/palo.2015.014
Thrombolite in Early Ordovician Microbial Reef,South China: A Case Study of the Huanghuachang Section in Yichang
TANG Haobo
(School of Earth Science and Resources, China University of Geosciences (Beijing) , Beijing 100083, China)
Abstract: A set of microbial reef is developed in the Honghuayuan Formation at the Huanghuachang section in the Yichang, Hubei Province. The microbial reef contains well-preserved thrombolites, one of the important types of microbial carbonate (i. e. thrombolite) . Based on the previous studies on the reef-building role of the metazoan and metazoan communities in Early Ordovician reefs in South China, this paper mainly studied the formation mechanism of thrombolite in microbial reefs by combining the macroscopic and microscopic characteristics of microbial reef through fieldwork and sampling and indoor mill observation. The results showed that the microbial reefs of the Honghuayuan Formation in the Huanghuachang section are dominated by thrombolite, accompanied by a small number of metazoan fossils, such as Calathium , lithistid sponge, bryozoan, etc. This reef represents a special type of microbial reefs in the Early Ordovician, that is, the microbial reefs are dominated by microbial carbonate, and locally metazoan is involved in the reef formation which is a special type of reef in the transition process from microbial reefs to metazoan reefs. At the same time, a large number of calcified cyanobacterial fossils are found in the thrombolite, such as Gerwanella, Subtifloria and Nuia, and the more intuitive calcified cyanobacteria showed that the thrombolites are the products of sedimentation, aggregation and early petrification of biofilm or mats dominated by cyanobacteria, indicating that there was a very close relationship between thrombolite and the complex calcification of microbial mats dominated by cyanobacteria. This study provides a typical example for the study of the formation mechanism of thrombolite in the Early Ordovician microbial reefs in South China and the transition period from microbial reefs to metazoan reefs.
Keywords: Early Ordovician, Microbial Reef, Thrombolite, Calcified Cyanobacteria Fossils, Huanghuachang Section, South China Plate
DOI: 10.48014/cesr.20241011001
Citation: TANG Haobo. Thrombolite in Early Ordovician microbial reef, South China: A case study of the Huanghuachang section in Yichang[J]. Chinese Earth Sciences Review, 2024, 3(4): 196-206.
